细胞松弛素杂聚体(meroaspochalasins,mAPOs)是丝状真菌黄柄曲霉产生的标志性代谢产物,因其迷人的化学结构以及抗肿瘤药物开发潜力,在天然产物和合成化学界引起了极大的兴趣。虽然细胞松弛素骨架生物合成的核心基因和部分修饰基因已经成功得到表征,但这些单体在微生物体内如何组装形成mAPOs仍然是个谜团。此外,野生黄柄曲霉中mAPOs的含量低且产率不稳定,而化学合成的方法步骤繁多,这严重限制了它们的进一步研究和开发。

近日,药学院张勇慧教授团队在国际权威期刊Angewandte Chemie International Edition发表了题为“Cooperative Redox Reactions Encoded by Two Gene Clusters EnableIntermolecular Cycloaddition Cascade for the Formation of Meroaspochalasins”的研究论文。本研究通过基因敲除、体外酶反应、蛋白结构预测和氨基酸定点突变,以及18O同位素标记等实验,阐明了聚合反应的两个关键中间体aspochalasinB(7)和聚酮双烯体前体化合物21(或26),以及终产物mAPOs的生物合成机制。
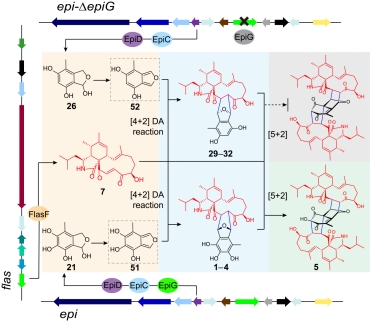
在本项研究中,作者在丝状真菌黄柄曲霉中鉴定了两条次生代谢产物生物合成基因簇,flas和epi,它们分别负责mAPOs中的两个亚单元,亲双烯体7和双烯体前体异苯骈半缩醛(21或26)的生物合成。其中,flas簇内的BBE-like氧化酶FlasF催化多种细胞松弛素的C18位羟基的氧化,生成相应的C18羰基细胞松弛素单体,其中就包括杂聚反应过程中关键的中间体7。此外,FlasF还可与短链脱氢酶/还原酶(SDR)FlasG,以及内源NADPH交互作用,进一步增加细胞松弛素单体合成代谢网络的复杂性。
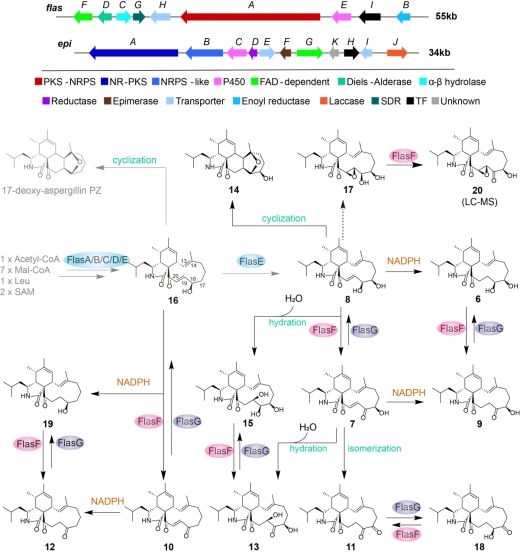
图1.flas和epi基因簇信息及细胞松弛素单体变换网络
在另一条基因簇epi中,细胞色素P450酶EpiC和SDR酶EpiD协同催化双烯体前体21(或26)的形成,随后它们自发脱水得到双烯体51(或52),在这一过程中,EpiD起着关键的“救援者”作用,防止聚酮C8位羟基的过度氧化和副产物内酯衍生物的形成。这一还原步骤确保了生物合成途径的适当调节,突出了EpiD作为控制双烯体形成过程中氧化和还原微妙平衡的重要作用。
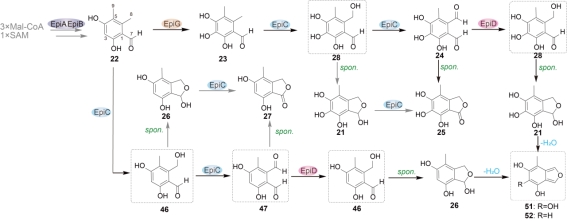
图2.双烯体51和52的生物合成通路
最后,亲双烯体7与双烯体51(或52)之间,可以通过级联的非酶促[4π + 2π] Diels-Alder反应和[5π + 2π]环加成反应,逐步产生mAPOs二聚体和三聚体。而epi簇中的黄素依赖氧化酶EpiG催化双烯体C3位的羟基化,这是形成mAPOs三聚体的关键步骤。
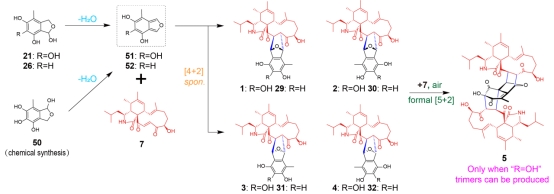
图3.细胞松弛素杂聚体(mAPOs)非酶聚合过程的阐明
综上所述,本文通过体内基因敲除、异源表达、体外生化表征、18O同位素标记以及相关蛋白计算等研究的结合,揭示了黄柄曲霉产生细胞松弛素异二聚体和三聚体的复杂、协作的生物合成机制。细胞松弛素杂聚体(mAPOs)已被本项工作证实是来源于两个独立BGC的跨基因簇DA环加成产物,并且其中两个独立单元的聚合被证明是非酶的过程。细胞松弛素杂聚体生物合成机制的成功阐明,不仅为这一过程建立了新的范式,也为工程高效生物合成细胞松弛素杂聚体奠定了基础,并为通过组合生物合成方式扩大结构多样性的策略开辟了新的途径。
药学院2021级博士研究生李鹏坤、2022级硕士研究生孟洁和博士后张晓天为论文共同第一作者,药学院张勇慧教授、周渊副教授、叶英副教授以及朱虎成教授为论文共同通讯作者。中国农业科学院深圳农业基因组研究所博士后张晓鹏、南京农业大学叶永浩教授、中国科学院青岛生物能源与过程研究所黄雪年研究员、德国马尔堡大学李书明教授也参与了本项研究。该项工作得到国家重点研发计划、国家自然科学基金等项目的支持。
原文链接:https://onlinelibrary.wiley.com/doi/full/10.1002/anie.202502766



